适应性免疫系统的多样性和分化
介绍
T 细胞受体 (TCR) 和 B 细胞受体 (BCR) 表现出惊人的多样性。虽然估计数字相差很大,但理论上可以有多达 10 25种不同的受体组合。在任何特定时刻,人体内可能都有大约 10 8种不同的受体。这种多样性仅仅来自七种不同的组成部分。单独来看,这些称为链的不同组成部分只能解释一小部分多样性:T 细胞的 alpha/beta 或 delta/gamma,B 细胞的重链/κ 或重链/λ。淋巴细胞多样性的关键在于链的编码方式。每条链由多个片段组成,每个片段由多个基因编码。这些片段重新组合产生大量不同的配置。
有关 BCR 和 TCR 的背景信息,请参阅我们的 T 细胞和 B 细胞概述页面
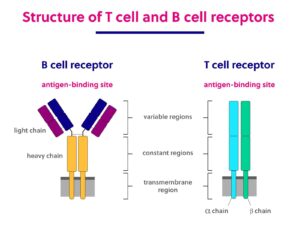
T 和 B 淋巴细胞的多种抗原受体
B 细胞和 T 细胞受体之间的多样性主要通过V(D)J 重组产生,这涉及可变区、多样性区、连接区和恒定区(分别缩写为 V、D、J 和 C)基因片段的改组和连接。为了简要介绍这些基因片段提供的变异潜力,下面描述了它们相对于 BCR 的分布和贡献:
- V:可变 (V) 基因片段编码轻链和重链的组成部分。有 29 到 46 个不同的 V 片段可以编码每种链类型。
- D:多样性(D)基因片段仅编码重链基因,每种重链类型有 23 个不同的 D 基因片段。
- J:连接(J)基因片段通过每种链类型 4-6 个不同的基因片段编码轻链和重链成分。
- C:恒定(C)基因片段通过每种链类型的 1-9 个不同片段编码轻链和重链成分。
在单个细胞中,V、D 和 J 片段各占一个。这些片段的独特组合决定了 B 细胞分泌的免疫球蛋白/抗体和 T 细胞受体的结合特异性和下游应用。
在细胞成熟的早期阶段,B 细胞的 V(D)J 重组发生在骨髓中,而 T 细胞的 V(D)J 重组发生在胸腺中。对于 B 细胞重链受体,一个 D 基因片段与一个 J 基因片段连接。然后,一个 V 基因片段添加到新形成的 DJ 复合体中。对于 T 细胞,这种 DJ 连接和 V 基因添加发生在 beta 链或 delta 链中。对于 B 细胞轻链受体和 T 细胞 alpha 或 gamma 链受体,一个 V 片段与一个 J 片段连接,不存在 D 基因。
有关 BCR 和 TCR 多样性与健康关系的信息,请参阅我们的免疫组库和适应组页面
可变区的结构和多样性
受体的重排 V(D)J 部分称为可变区或 V 区,因其负责抗原结合和特异性而备受关注。V 区由框架 (FR) 和互补决定区 (CDR) 组成。当 V(D)JC 翻译成氨基酸序列时,V 区可进一步细分为几个部分,包括前导序列、FR1、CDR1、FR2、CDR2、FR3、CDR3、FR4 和 C 域。C 域编码各种恒定区。
CDR3 尤其令人感兴趣,因为它是抗原结合位点中变化最大的部分,一些研究表明 CDR3 与抗原特异性特别相关。CDR3 区域横跨 V(D)J 连接。CDR3 区域中 D 段的两侧是可变的“N”区域。N 区域是通过删除几个碱基然后添加随机序列生成的,因此具有高度可变性。

体细胞重组、等位基因排除和克隆扩增
上面描述的 V(D)J 重组涉及 体细胞重组,这种重组发生在幼稚、失活的细胞中。以前人们认为,单个 B 细胞或 T 细胞产生的所有受体对于该细胞都是相同的。 等位基因排除这个术语定义的是,尽管每个细胞中都有两个重组 V(D)JC 区域,但只有一个副本表达。然而,有新的证据表明, 等位基因包含 也可以发生,尽管比例很低,这破坏了以前一个重组受体对应一个细胞化学计量的信念。当幼稚 B 细胞或 T 细胞的受体识别抗原时,细胞就会被激活,然后增殖,这一过程称为克隆扩增。这种激活和扩增过程正是免疫系统产生特异性适应性反应的原因。
B 细胞成熟
在激活和克隆扩增之后,B 细胞会经历成熟过程,通常分为三个步骤:1) 同种型转换、2) 体细胞超突变和 3) 亲和力成熟。不过,这些步骤的顺序各不相同,有些细胞可能会完全跳过一个或多个步骤。
同型转换
同种型转换,也称为类别转换重组,可增强 B 细胞分泌抗体的效应功能。抗体重链和 BCR 分为不同的同种型(IgM、IgD、IgG、IgA 或 IgE),横跨可变区和恒定区。可变区识别抗原,而恒定区尾部(即可结晶片段 (Fc) 区)与补体系统或细胞表面受体中的蛋白质相互作用。
幼稚 B 细胞首先表达 IgM 和/或 IgD 同型,直至活化,从而转换为 IgA、IgE 或 IgG 同型。每种同型均可对免疫反应产生不同的影响,包括诱导抗体依赖性细胞介导的细胞毒性 (ADCC) 或抗体依赖性细胞介导的吞噬作用 (ACDP)、肥大细胞或嗜碱性粒细胞脱粒和/或激活补体系统。
BCR 和抗体同型由激活 BCR、辅助受体(例如 CD40)、细胞因子受体和 Toll 样受体 (TLR) 获得的不同信号组合决定。这些信号将激活活化诱导胞苷脱氨酶 (AID) 酶,从而诱发错配突变,随后发生双链断裂。这些断裂通过重链“转换区”中的非同源末端连接 (NHEJ) 修复。重链的恒定区由不同的 C H外显子组成,每个外显子前面都有一个转换区和内含子启动子。根据激活 AID 的信号,特定的 C H外显子将被切除,剩余的 C H外显子组合将决定 BCR 同型。
总之,同种型转换允许针对同一靶抗原产生不同的免疫效应子功能。由于同种型类别由恒定区决定,因此使用从高度可变的 CDR3 区延伸到恒定区的免疫测序技术对于更全面地分析 B 细胞抗原结合和同种型转换非常重要。
体细胞超突变和亲和力成熟
体细胞超突变和亲和力成熟在某种程度上是相互关联的。在体细胞超突变中,BCR 基因位点随机突变以产生额外的多样性。这些突变可以增加、减少或不影响抗原结合亲和力。通常,由于体细胞超突变而具有增加亲和力的 B 细胞比没有增加亲和力的 B 细胞增殖得更快。这种突变和增殖以产生一组对其同源抗原具有更高结合亲和力的 B 细胞的过程称为亲和力成熟。
有关 BCR 和 TCR 多样性与健康关系的信息,请参阅我们的免疫组库和适应组页面